
Węzeł zatokowo-przedsionkowy - Sinoatrial node
| Węzeł zatokowo-przedsionkowy | |
|---|---|
 Węzeł zatokowo-przedsionkowy pokazany jako 1) reszta układu przewodzącego serca w zestawie pokazana na niebiesko.
| |
| Detale | |
| System | System przewodzenia elektrycznego serca |
| Tętnica | Tętnica węzłowa zatokowo-przedsionkowa |
| Identyfikatory | |
| łacina | nodus sinuatrialis |
| Akronim(y) | węzeł SA |
| Siatka | D012849 |
| TA98 | A12.1.06.003 |
| TA2 | 3953 |
| FMA | 9477 |
| Terminologia anatomiczna | |
Węzeł zatokowo-przedsionkowy (znany również jako sinuatrial węzeł , węzeł SA lub węzła zatokowego ) to grupa komórek zwanych komórkami stymulatora , znajdujących się w ścianie prawego przedsionka w sercu . Komórki te mają zdolność spontanicznego wytwarzania impulsu elektrycznego ( potencjału czynnościowego ), który przemieszcza się przez system przewodzenia elektrycznego serca powodując jego skurcz . W zdrowym sercu węzeł SA nieprzerwanie wytwarza potencjały czynnościowe, ustalając rytm serca ( rytm zatokowy ) i dlatego jest znany jako naturalny rozrusznik serca . Na tempo wytwarzanych potencjałów czynnościowych (a tym samym częstość akcji serca ) wpływają nerwy, które je dostarczają.
Struktura
Węzeł zatokowo-przedsionkowy jest strukturą w kształcie banana o różnej wielkości, zwykle o długości 10-30 milimetrów (mm), szerokości 5-7 mm i głębokości 1-2 mm.
Lokalizacja
Węzeł SA zlokalizowany jest w ścianie ( nasierdziu ) prawego przedsionka , bocznie od wejścia do żyły głównej górnej w rejonie zwanym zatoką venarum (stąd sino- + przedsionek ) . Znajduje się mniej więcej pomiędzy rowkiem zwanym crista terminals, znajdującym się na wewnętrznej powierzchni serca, a odpowiadającym mu bruzdą końcową , na powierzchni zewnętrznej. Rowki te biegną między wejściem żyły głównej górnej i żyły głównej dolnej .
Mikroanatomia

Komórki węzła SA są rozrzucone w siatce tkanki łącznej zawierającej nerwy , naczynia krwionośne , kolagen i tłuszcz . Bezpośrednio otaczające komórki węzła SA są komórkami paranodalnymi. Komórki te mają struktury pośrednie między komórkami węzła SA a resztą przedsionka . Tkanka łączna wraz z komórkami paranodnymi izoluje węzeł SA od reszty przedsionka, zapobiegając wpływowi aktywności elektrycznej komórek przedsionkowych na komórki węzła SA. Komórki węzła SA są mniejsze i jaśniejsze niż otaczające komórki przedsionkowe , przy czym średnia komórka ma około 8 mikrometrów średnicy i 20-30 mikrometrów długości (1 mikrometr = 0,000001 metra). W przeciwieństwie do komórek przedsionkowych, komórki węzła SA zawierają mniej mitochondriów i włókien mięśniowych , a także mniejszą siateczkę sarkoplazmatyczną . Oznacza to, że komórki węzła SA są mniej przystosowane do kurczenia się w porównaniu z komórkami przedsionków i komór .
Potencjały czynnościowe przechodzą z jednej komórki serca do drugiej przez pory znane jako połączenia szczelinowe. Te połączenia szczelinowe są zbudowane z białek zwanych koneksynami . W węźle SA jest mniej połączeń szczelinowych i są one mniejsze. Jest to ponownie ważne w izolowaniu węzła SA od otaczających komórek przedsionkowych.
Dopływ krwi
Węzeł zatokowo-przedsionkowy otrzymuje dopływ krwi z tętnicy zatokowo - przedsionkowej . Jednak ten dopływ krwi może się znacznie różnić między osobnikami. Na przykład u większości ludzi jest to pojedyncza tętnica , chociaż w niektórych przypadkach do węzła SA zaopatrują się 2 lub 3 tętnice węzła zatokowo-przedsionkowego. Ponadto tętnica węzła SA pochodzi głównie z odgałęzienia prawej tętnicy wieńcowej ; jednak u niektórych osób wywodzi się z tętnicy okalającej , która jest odgałęzieniem lewej tętnicy wieńcowej . Wreszcie, tętnica węzła SA zwykle przechodzi za żyłą główną górną , zanim dotrze do węzła SA; jednak w niektórych przypadkach przechodzi z przodu. Pomimo tych wielu różnic, wydaje się, że nie ma żadnej korzyści w stosunku do liczby tętnic węzła zatokowo-przedsionkowego danej osoby ani ich pochodzenia
Drenaż żylny
Nie ma dużych żył, które odprowadzają krew z węzła SA. Zamiast tego mniejsze żyłki odprowadzają krew bezpośrednio do prawego przedsionka .
Funkcjonować
Rozpalanie
Główną rolą komórki węzła zatokowo-przedsionkowego jest inicjowanie potencjałów czynnościowych serca, które mogą przechodzić przez komórki mięśnia sercowego i powodować skurcz. Potencjał czynnościowy to szybka zmiana potencjału błonowego , wytworzona przez ruch naładowanych atomów ( jonów ). W przypadku braku stymulacji komórki nierozrusznikowe (w tym komórki komorowe i przedsionkowe ) mają względnie stały potencjał błonowy; nazywa się to potencjałem spoczynkowym . Ta faza spoczynku (patrz potencjał czynnościowy serca, faza 4 ) kończy się, gdy potencjał czynnościowy dociera do komórki. Powoduje to pozytywną zmianę potencjału błony, znaną jako depolaryzacja , która rozprzestrzenia się w sercu i inicjuje skurcze mięśni . Komórki rozrusznika nie mają jednak potencjału spoczynkowego. Zamiast tego, natychmiast po repolaryzacji , potencjał błonowy tych komórek zaczyna ponownie automatycznie depolaryzować, zjawisko znane jako potencjał stymulatora . Gdy potencjał stymulatora osiągnie ustawioną wartość, potencjał progowy , wytwarza potencjał czynnościowy. Inne komórki w sercu (w tym włókna Purkiniego i węzeł przedsionkowo-komorowy ) mogą również inicjować potencjały czynnościowe; jednak robią to wolniej, a zatem, jeśli węzeł SA działa prawidłowo, jego potencjały czynnościowe zwykle przewyższają te, które byłyby wytwarzane przez inne tkanki.
Poniżej przedstawiono 3 fazy potencjału czynnościowego węzła zatokowo-przedsionkowego. W potencjale czynnościowym serca występuje 5 faz (oznaczonych 0-4), jednak potencjały czynnościowe stymulatora nie mają oczywistej fazy 1 lub 2.
Faza 4

Ta faza jest również znana jako potencjał stymulatora . Bezpośrednio po repolaryzacji, gdy potencjał błony jest bardzo ujemny (jest hiperpolaryzowany), napięcie powoli zaczyna rosnąć. Jest to początkowo spowodowane zamknięciem kanałów potasowych , co zmniejsza wypływ jonów potasu (I k ) z komórki (patrz faza 2 poniżej). Hiperpolaryzacja powoduje również aktywację aktywowanych hiperpolaryzacją kanałów cyklicznych bramkowanych przez nukleotydy (HCN) . Aktywacja kanałów jonowych przy bardzo ujemnych potencjałach błonowych jest niezwykła, dlatego przepływ sodu (Na + ) i trochę K + przez aktywowany kanał HCN określany jest jako prąd śmieszny (I f ). Ten zabawny prąd powoduje, że potencjał błonowy komórki stopniowo się zwiększa, ponieważ ładunek dodatni (Na + i K + ) wpływa do komórki. Innym mechanizmem zaangażowanym w potencjał stymulatora jest zegar wapniowy . Odnosi się to do spontanicznego uwalniania wapnia z retikulum sarkoplazmatycznego (magazynu wapnia) do cytoplazmy, znanego również jako iskry wapniowe . Ten wzrost wapnia w komórce aktywuje następnie wymiennik sód-wapń (NCX), który usuwa jeden Ca 2+ z komórki i wymienia go na 3 Na + w komórce (a tym samym usuwa ładunek +2 z komórki, ale pozwalając, aby ładunek +3 dostał się do komórki) dalej zwiększając potencjał błonowy. Wapń później ponownie wchodzi do komórki przez SERCA i kanały wapniowe znajdujące się na błonie komórkowej. Wzrost potencjału błonowego wytworzony przez te mechanizmy aktywuje kanały wapniowe typu T, a następnie kanały wapniowe typu L (które otwierają się bardzo powoli). Kanały te umożliwiają przepływ Ca 2+ do komórki, dzięki czemu potencjał błonowy jest jeszcze bardziej dodatni.
Faza 0
To jest faza depolaryzacji. Gdy potencjał błonowy osiąga potencjał progowy (około -20 do -50 mV), komórka zaczyna gwałtownie depolaryzować (stać się bardziej dodatnia). Wynika to głównie z przepływu Ca 2+ przez kanały wapniowe typu L, które są teraz całkowicie otwarte. Na tym etapie kanały wapniowe typu T i kanały HCN ulegają dezaktywacji.
Faza 3
Ta faza jest fazą repolaryzacji. Dzieje się tak z powodu dezaktywacji kanałów wapniowych typu L (zapobiegając przemieszczaniu się Ca 2+ do komórki) oraz aktywacji kanałów potasowych, co umożliwia wypływ K + z komórki, przez co potencjał błonowy jest bardziej ujemny.
Zaopatrzenie nerwów
Częstość akcji serca zależy od szybkości, z jaką węzeł zatokowo-przedsionkowy wytwarza potencjały czynnościowe . W spoczynku tętno wynosi od 60 do 100 uderzeń na minutę. Jest to wynikiem działania dwóch zestawów nerwów, jednego działającego w celu spowolnienia wytwarzania potencjału czynnościowego (są to nerwy przywspółczulne ), a drugiego działającego w celu przyspieszenia wytwarzania potencjału czynnościowego ( nerwy współczulne ).
Nerwy współczulne zaczynają się w odcinku piersiowym rdzenia kręgowego (w szczególności T1-T4). Nerwy te uwalniają neuroprzekaźnik zwany noradrenaliną (NA) . Wiąże się to z receptorem na błonie węzła SA, zwanym beta-1adrenoceptorem . Wiązanie na to ten receptor uaktywnia białkiem G (w szczególności G S -Białko S dla stymulacyjnej), który inicjuje cykl reakcji (tzw szlaku cAMP ), który prowadzi do wytwarzania cząsteczki zwany cykliczny adenosinemonophosphate (cAMP ) . Ten cAMP wiąże się z kanałem HCN (patrz wyżej). Wiązanie cAMP z HCN zwiększa przepływ Na + i K + do komórki, przyspieszając potencjał rozrusznika, a więc wytwarzając potencjały czynnościowe w szybszym tempie i zwiększając częstość akcji serca. Wzrost częstości akcji serca jest znany jako pozytywna chronotropia .
W przywspółczulne nerwów zaopatrujących węzeł SA (w szczególności nerwy vagus ) pochodzą z mózgu . Nerwy te uwalniają neuroprzekaźnik zwany acetylocholiną (ACh) . ACh wiąże się z receptorem zwanym receptorem muskarynowym M2 , zlokalizowanym na błonie węzła SA. Aktywacja tego receptora M2, aktywuje białko zwane białkiem G (zwłaszcza G ı białka , i dla hamowania). Aktywacja tego białka G blokuje szlak cAMP, zmniejszając jego działanie, a tym samym hamując aktywność współczulną i spowalniając wytwarzanie potencjału czynnościowego. Oprócz tego białko G aktywuje również kanał potasowy, który pozwala K + wypływać z komórki, co powoduje, że potencjał błonowy jest bardziej ujemny i spowalnia potencjał rozrusznika, zmniejszając w ten sposób tempo wytwarzania potencjału czynnościowego, a tym samym zmniejszając pracę serca wskaźnik. Spadek częstości akcji serca jest znany jako ujemna chronotropia .
Pierwsza komórka wytwarzająca potencjał czynnościowy w węźle SA nie zawsze jest taka sama: jest to znane jako przesunięcie stymulatora. U niektórych gatunków zwierząt – na przykład u psów – przesunięcie wyższe (tzn. komórka, która wytwarza najszybszy potencjał czynnościowy w węźle SA jest wyższy niż poprzednio) zwykle powoduje zwiększenie częstości akcji serca, podczas gdy przesunięcie gorsze (tzn. komórka wytwarzająca najszybszy potencjał czynnościowy w węźle SA jest niższy niż poprzednio) powoduje zmniejszenie częstości akcji serca.
Znaczenie kliniczne
Dysfunkcja węzła zatokowego opisuje nieregularne bicie serca spowodowane wadliwymi sygnałami elektrycznymi serca. Kiedy węzeł zatokowo-przedsionkowy serca jest uszkodzony, rytmy serca stają się nieprawidłowe – zwykle zbyt wolne lub wykazujące przerwy w jego funkcji lub kombinacji i bardzo rzadko szybsze niż normalnie.
Zablokowanie dopływu krwi tętniczej do węzła SA (najczęściej z powodu zawału mięśnia sercowego lub postępującej choroby wieńcowej ) może zatem powodować niedokrwienie i śmierć komórek w węźle SA. Może to zakłócić funkcję elektrycznego stymulatora węzła SA i spowodować zespół chorej zatoki .
Jeśli węzeł SA nie działa lub impuls generowany w węźle SA zostanie zablokowany, zanim przejdzie przez system przewodzenia elektrycznego, grupa komórek w głębi serca stanie się jego rozrusznikiem.
Historia
Węzeł zatokowo-przedsionkowy został po raz pierwszy odkryty przez młodego studenta medycyny, Martina Flacka , w sercu kreta , podczas gdy jego mentor, Sir Arthur Keith , jechał z żoną na rowerze. Oni dokonali odkrycia w laboratorium prowizorycznym ustanowionego na farmie w hrabstwie Kent , w Anglii , o nazwie Manna Place. Ich odkrycie zostało opublikowane w 1907 roku.
Dodatkowe obrazy

Serce; system przewodzenia (węzeł SA oznaczony 1)
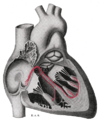
Schematyczne przedstawienie pęczka przedsionkowo-komorowego

Zobacz też
Bibliografia
Zewnętrzne linki
- Figura anatomiczna: 20:06-01 w Human Anatomy Online, SUNY Downstate Medical Center – „System przewodzenia serca”.
- Schemat na gru.net
- thoraxlesson4 na lekcji anatomii Wesleya Normana (Georgetown University) ( thoraxheartinternalner )
- https://web.archive.org/web/20070929080346/http://www.healthyheart.nhs.uk/heart_works/heart03.shtml
{kind=link}
{kind=link}